
MALE REPRODUCTIVE PHYSIOLOGY
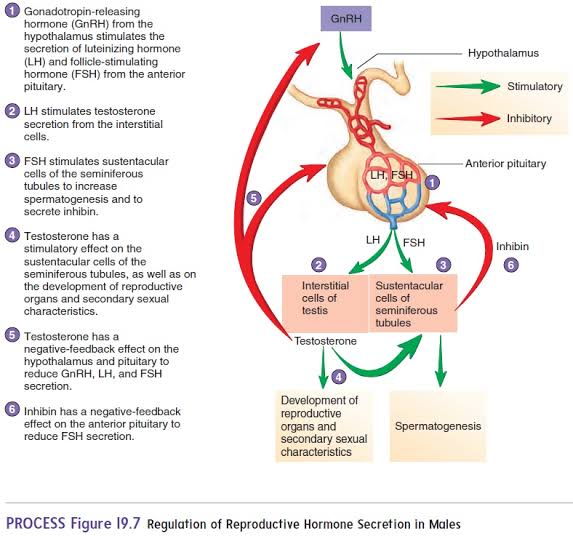
All aspects of male reproductive physiologyare under the endocrine control of the two major gonadotrophins, luteinizing hormone(LH) and follicle-stimulating hormone (FSH). The secretion of LH is pulsatile, with irregular episodes of secretion occurring every 2–4 hours. The actions of LH are primarily upon the Leydig cell, where, acting through adenylate cyclase, it promotes steroidogenesisby regulating the rate-limiting step of steroidogenesis; namely, conversion of cholesterol into the testosterone precursor, pregnenolone. Peak testosterone concentrations follow those of LH by about 40 minutes and decline back to prestimulation values over a further 40–80 minutes. Testosterone is required for the production of sperm in the testis and their subsequent maturation in the epididymis, for the function of the accessory sex glands and for the development of masculine secondary sexual characteristics. After aromatization into oestrogen within the brain, testosterone is also responsible for negative-feedback regulation of LH secretionand for male behaviour. Curiously, in long-term castrated animals, neither negative feedback nor libido can be restored by testosterone administration, for brain aromatase activity is eventually lost, and oestrogen itself has to be given for the restoration of these effects. Within the lumen of the seminiferous tubule, testosterone is converted by 5-reduction into 5-dihydrotestosterone (DHT), which is not susceptible to aromatization and is a more potent androgen than testosterone itself. Both testosterone and DHT are bound within the tubule lumen by the secretory product of the Sertoli cells, androgen-binding protein (ABP). The role of ABP therefore appears to be to maintain high androgen concentrations in the lumina of the seminiferous tubule and epididymis.
The main target of FSH is the Sertoli cell, where it also acts through adenylate cyclase-linked enzyme systems. Under the influence of FSH, Sertoli cells secrete ABP and aromatize testosterone into oestrogens. Adeqate FSH stimulation is also required to permit Sertoli cells to support spermatogenesis. Some evidence suggests that the production of pyruvate and lactate, which act as energy substrates for germ cells, may be a key role of the FSH-stimulated activity of the Sertoli cell in maintaining spermatogenesis. Debate remains over the pattern of secretion of FSH; some consider it to be pulsatile, a manner analogous to LH, while others consider its secretion only to exhibit longer-term fluctuations. FSH secretion is regulated by both gonadal steroids and inhibin, the regulatory protein secreted by Sertoli cells
The actions of the gonadotrophins are, however, not limited to the somatic cells of the testis, as both LH and FSH regulate aspects of germ cell activity. For example, experiments upon hypophysectomized rams indicated that the rate of division of stem spermatogonia is controlled by LH, while the rate of subsequent divisions and the ability of cells to undergo meiosis are regulated by FSH. Whilst these actions upon germ cells are, in part, mediated through the stimulation of activity in the somatic component of the testis by the gonadotrophins, circumstantial evidence indicates direct actions upon germ cells themselves.
At the end of the lesson students should learn about
1. spermatogenesis
2. Functions of male sex hormones


Download Files
- Chapters 4
- Department Physiology
- Teacher
Dr. Faryal Mazhar
